
Spermidine reduces cancer-related mortality in humans
Spermidine, a polyamine that is particularly abundant in sperm, is a polycation that is associated with DNA in most, if not all, living organisms. If in excess over DNA, spermidine is volatile and confers the characteristic smell and taste to sperm. Spermidine is contained in all food items containing nuclei (such as vegetable, meat and fish), is scarcely present in milk and hyperprocessed food items, yet is overabundant in food products generated by bacterial or fungal fermentation (such as smelly mature cheese and the soya fermentation product natto) or specific fruits with a sperm-like odor (such as durian fruit) A recent prospective, observational epidemiological study revealed that individuals that have been eating a diet that is spermidine-rich are characterized by a reduced overall mortality, as well as a decreased mortality by each of the major causes of death, namely (i) cardiovascular, (ii) cancer and (iii) ‘other’ causes [1]. The association between high spermidine uptake and reduced mortality is independent from confounding factors including age, sex, body mass index, consumption of alcohol or aspirin, metabolic syndrome, diabetes, physical activity, and socioeconomic status, as well as conventional dietary scores distinguishing healthy from unhealthy eating. These results confirm our long-lasting suspicion that spermidine has broad health-improving effects [2–9], based on our observation that nutritional supplementation of this polyamine can extend the longevity of multiple model species, including yeast, nematodes, flies and mice [10–12]. Here, we explore the literature linking spermidine to reduced cancer risk.
Spermidine, a polyamine that is particularly abundant in sperm, is a polycation that is associated with DNA in most, if not all, living organisms. If in excess over DNA, spermidine is volatile and confers the characteristic smell and taste to sperm. Spermidine is contained in all food items containing nuclei (such as vegetable, meat and fish), is scarcely present in milk and hyperprocessed food items, yet is overabundant in food products generated by bacterial or fungal fermentation (such as smelly mature cheese and the soya fermentation product natto) or specific fruits with a sperm-like odor (such as durian fruit) A recent prospective, observational epidemiological study revealed that individuals that have been eating a diet that is spermidine-rich are characterized by a reduced overall mortality, as well as a decreased mortality by each of the major causes of death, namely (i) cardiovascular, (ii) cancer and (iii) ‘other’ causes [1]. The association between high spermidine uptake and reduced mortality is independent from confounding factors including age, sex, body mass index, consumption of alcohol or aspirin, metabolic syndrome, diabetes, physical activity, and socioeconomic status, as well as conventional dietary scores distinguishing healthy from unhealthy eating. These results confirm our long-lasting suspicion that spermidine has broad health-improving effects [2–9], based on our observation that nutritional supplementation of this polyamine can extend the longevity of multiple model species, including yeast, nematodes, flies and mice [10–12]. Here, we explore the literature linking spermidine to reduced cancer risk.
Beyond its metabolic function as an intermediate in the conversion of putrescine to spermine, and it role as a precursor of acetyl-spermidine [13,14], spermidine functions as a signaling molecule that acts as an endogenous inhibitor of the acetyl transferase EP300, an enzyme that transfers acetyl groups from acetyl coenzyme A (CoA) on lysine residues of cytoplasmic and nuclear proteins (Figure 1(a)) [15]. This inhibitory effect results from the sterical competition between spermidine and acetyl CoA for binding to the EP300 catalytic site [12]. Acetyl CoA functions as an endogenous inhibitor of macroautophagy/autophagy, meaning that excess acetyl CoA (and in particular its cytosolic pool) cause the hyperacetylation of multiple proteins involved in the regulation or execution of autophagy, thus stalling the process [16–18]. This may constitute a phylogenetically ancient mechanism through which nutrient excess, which results in an increase in cellular acetyl CoA, inhibits autophagy [19,20]. Conversely, fasting or caloric restriction cause a reduction in acetyl CoA levels, thereby triggering autophagy [21]. This ancient regulatory system (conserved throughout eukaryotic evolution) facilitates the adaptation of organisms to dwindling nutrient resources. Intriguingly, this regulatory system may be modulated by endogenous metabolites such as spermidine, as well as external agents such as aspirin. Aspirin, which – like spermidine – inhibits EP300 by competing with acetyl CoA [22], is well known for its cancer-preventive action [23,24] in particular on the gastrointestinal system (a 30% decrease of cancer in long-term users) [25] but also lung [26], mammary [27] and prostate carcinomas (a 10% decrease) [28].
It is hence tempting to speculate, yet remains to be demonstrated, that both spermidine and aspirin share a common mode of action with respect to the prophylaxis of malignant disease. As mentioned above, spermidine is a potent inducer of autophagy, both in vitro (in cultured cells) and in vivo (when spermidine is supplied with the food or drinking water). In yeast, nematodes and flies, life span extension by spermidine is lost upon knockout/knockdown of essential autophagy-related (ATG) genes (such as ATG5, ATG7 or BECN1) [10]. Similarly, in mice, the capacity of spermidine to prevent cardiac aging [29] is fully lost upon cardiomyocyte-specific knockout of Atg7 [11,15]. These findings suggest that the desirable effects of spermidine on health are secondary to the induction of autophagy. In a model of cancer prophylaxis using hydroxycitrate, an inhibitor of the acetyl CoA-generating enzyme ACLY (ATP citrate lyase), which reduces intracellular acetyl CoA levels, we observed that inactivation of ATG5 in KRAS-expressing non-small cell lung cancer (NSCLC) cells abolishes the chemopreventive effect [30,31]. Similarly, both hydroxycitrate and spermidine could enhance the anticancer immune response induced by immunogenic chemotherapy, thus improving the control of established tumors. Again, this effect is lost upon knockdown of essential autophagy genes (ATG5 or ATG7) [31,32], supporting the idea that spermidine might induce autophagy in cancer cells to improve anticancer immunosurveillance.
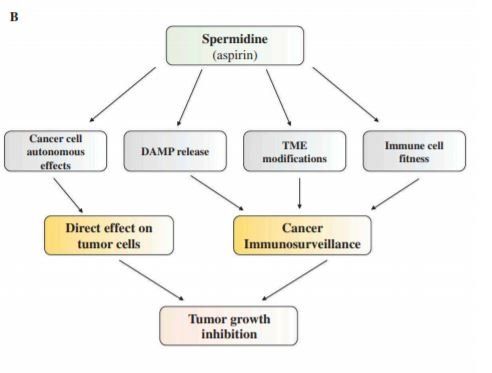
Along the same line of evidences, the activation of autophagy elicited by spermidine administration accounts for the prevention of hepatocellular carcinoma formation [33], reduces the burden of colorectal cancer allografts [34] and promotes the rejuvenation of the adaptive branch of the immune system [35,36]. The history of tumor biology has been marked by a long period during which malignancy was thought to solely result from genetic and epigenetic aberrations in the cancer cells. However, it has become clear over the past decade that beyond such cellautonomous events, cancer will only emerge and develop into a life-threatening disease if the immune system fails to detect and eliminate malignant cells (Figure 1(b)) [37–39]. Autophagy has classically been studied with respect to cancer, while adopting the cell-autonomous point of view, often in vitro and in xenotransplantation experiments (in which human cancer cells are implanted into immunodeficient hosts). Many of these studies came to the conclusion that autophagy can increase the fitness of malignant cells, acting as part of a cellular defense system against stressful or harmful conditions, while some investigators noted the capacity of autophagy to suppress malignant transformation in specific circumstances, for instance by inhibiting pro-carcinogenic inflammatory reactions [40–45]. However, it appears important to note that, in the presence of an immune system, the anticancer effects of autophagy induction within malignant cells prevail because of the effective induction of antitumor immunity [46–52].
It will be important to study the detailed mechanisms through which spermidine and aspirin can stimulate immunosurveillance and to develop strategies for replacing them with more effective EP300 inhibitors or combining them with other immunostimulants.
Acknowledgments
GK is supported by the Ligue contre le Cancer (équipe labellisée); Agence National de la Recherche (ANR) – Projets blancs; ANR under the frame of E-Rare-2, the ERA-Net for Research on Rare Diseases; Association pour la recherche sur le cancer (ARC); Cancéropôle Ile-de-France; Chancelerie des universités de Paris (Legs Poix), Fondation pour la Recherche Médicale (FRM); a donation by Elior; the European Commission (ArtForce); European Research Area Network on Cardiovascular Diseases (ERA-CVD, MINOTAUR); the European Research Council (ERC); Fondation Carrefour; Institut National du Cancer (INCa); Inserm (HTE); Institut Universitaire de France; LeDucq Foundation; the LabEx Immuno-Oncology; the RHU Torino Lumière; the Seerave Foundation; the SIRIC Stratified Oncology Cell DNA Repair and Tumor Immune Elimination (SOCRATE); the SIRIC Cancer Research and Personalized Medicine (CARPEM); and the Paris Alliance of Cancer Research Institutes (PACRI). F.M. is grateful to the Austrian Science Fund FWF (Austria) for grants P23490-B20, P29262, P24381, P29203 P27893, I1000, “SFB Lipotox” (F3012), and DKplus Metabolic and Cardiovascular Diseases (W1226), as well as to Bundesministerium für Wissenschaft, Forschung und Wirtschaft and the Karl-Franzens University for grants “Unkonventionelle Forschung”. We acknowledge support from NAWI Graz and the BioTechMed-Graz flagship project “EPIAge.” FC is supported by the Ligue contre le Cancer.
Disclosure statement
Frank Madeo, Didac Carmona-Gutierrez, Oliver Kepp and Guido Kroemer are the scientific co-founders of Samsara Therapeutics. Frank Madeo and Didac Carmona-Gutierrez are scientific co-founders of Longevity Labs.
Funding
This work was supported by the Ligue contre le Cancer (équipe labellisée); Agence National de la Recherche (ANR); ERA-Net for Research on Rare Diseases; Association pour la recherche sur le cancer (ARC); Cancéropôle Ilede-France; Fondation pour la Recherche Médicale (FRM); European Commission (ArtForce); European Research Area Network on Cardiovascular Diseases (ERA-CVD, MINOTAUR); European Research Council (ERC); Fondation Carrefour; Institut National du Cancer (INCa); Inserm (HTE); Institut Universitaire de France; LeDucq Foundation; LabEx Immuno-Oncology; RHU Torino Lumière; the Seerave Foundation; SIRIC Stratified Oncology Cell DNA Repair and Tumor Immune Elimination (SOCRATE); SIRIC Cancer Research and Personalized Medicine (CARPEM); Paris Alliance of Cancer Research Institutes (PACRI); Austrian Science Fund FWF (Austria); [P23490-B20]; [P29262]; [P24381]; [P29203]; [P27893]; [I1000].
1.888.394.3394
1.307.218.9831
Sheridan, Wyoning, USA
www.FWGERX.com
ALL SALES FINAL
*These statements have not been evaluated by the Food and Drug Administration. These products are not intended to diagnose, treat, cure or prevent any disease.
Copyright © 2014 FWGE RX BioSCIENCE - All Rights Reserved.